 2021, Vol. 58
2021, Vol. 58



2. 西藏农牧学院高原生态研究所, 西藏林芝 860000
2. Institute of Plateau Ecology, Tibet College of Agriculture and Animal Husbandry, Linzhi, Tibet 860000, China
世界植被中,铁杉(Tsuga)是北美洲北部常绿针叶林的重要成分,而在同纬度的欧亚大陆,铁杉林仅有限地分布于海洋性气候所涉及的地域,在中国中纬度地区,铁杉林成为亚热带山地植被垂直带中的一个显著类型,尤其是青藏高原东南缘山地(包括川西滇西北),铁杉林占有十分显著的地位,成为我国植被的特点之一[1]。铁杉林是暗针叶林的类型之一,为亚热带地区所特有,是亚热带山地垂直带的代表类型[2]。
生长在西藏的暗针叶林主要由云杉属、冷杉属和铁杉属的树种组成,是欧亚大陆暗针叶林分布区的西南限界,分布于喜马拉雅山脉、念青唐古拉山脉以及横断山脉的湿润亚高山地带。西藏暗针叶林占全自治区森林总面积的48%、总蓄积量的61%,在高海拔森林生态系统中发挥着不可替代的重要作用。
迄今为止,针对西藏暗针叶林的云杉属和冷杉属开展的研究较多[2-7],对铁杉属的研究则明显不足[8],现有的研究成果主要集中在地上部分的植物群落[2-5],对地下生态系统所开展的调查研究工作则较少[6-7],目前尚未见有关西藏铁杉林土壤线虫方面的研究报道。
在森林生态系统中,土壤线虫占据着地下食物网的关键点位[9],是土壤生态系统的重要功能组分。土壤线虫通过与植物根系以及地下食物网中其他生物的相互作用,影响着生态系统的物质循环和能量流动,同时,地上植物群落也会影响到土壤生态系统物质和能量的输入,从而驱动土壤线虫群落的结构和功能发生相应的变化[10]。因此,土壤线虫群落兼具生物指示和影响生态系统的物质循环和能量流动功能组份等多重作用。
嘎隆拉山地处横断山与喜马拉雅山的交汇地带,高差显著,植被具有连续垂直分布的特点。本研究以西藏东南部具有代表性的墨脱嘎隆拉山铁杉林作为研究对象,目的是了解该地区铁杉林土壤线虫群落具有怎样的结构以及多样性特征,及其与环境因子之间的关系,同时,也试图利用土壤线虫群落的环境指示作用来分析该生态系统的稳定性状况及演替阶段,研究结果对于揭示藏东南地区铁杉林生态系统土壤线虫多样性规律,探讨地下与地上过程的相互作用和相互联系,认识土壤线虫群落在亚高山/高山森林土壤生态过程中的作用具有重要科学意义。
1 材料与方法 1.1 研究区概况嘎隆拉山位于西藏自治区林芝市波密县和墨脱县的交界处,是岗日嘎布山脉的西端段(也是横断山脉的西北端与喜马拉雅山交汇处),平均海拔4 800 m,年均气温10.4 ℃,年均降水量1 358 mm[11]。其南坡下延进入墨脱河谷,雅鲁藏布江向南拐弯后流经该河谷。由于雅鲁藏布江强烈下切,在短距离内出现巨大高差,因而该区域的垂直气候特点非常显著。来自印度洋的暖湿气流在嘎隆拉南坡与青藏高原低温气流交锋,导致该地气候多变,常年多雨雪。样地概况见表 1。样地1位于山脊,样地2、3设在相对平坦的山坡,样地4位于河边滩地,样地1为铁杉林(Tsuga dumosa)纯林,接近铁杉林在本区域分布的下限,样地2、3为铁杉和墨脱冷杉(Abies delavayi var. motuoensis)的混交林,样地4接近铁杉林在本区域分布的上限,为天然更新的幼树林。
|
|
表 1 样地概况 Table 1 Basic information of the sample plot relative to location |
2017年9月,按照植物群落和地形的差异分别设置50 m×20 m、50 m×20 m、87 m×20 m、10 m×10 m样地4个,使用内径7 cm土钻在每个样地内按随机布点方式多点(至少5点)分层采集土样,并按土层深度0~5 cm、5~10 cm、10~15 cm、15~0 cm、20~25 cm分别组成混合样。采集土壤样品共20个。将土样装入聚乙烯袋中,贴上标签后冷藏保存于装有冰袋的保温箱中带回实验室进行后续测定。
土壤理化性质测定[12]:土壤含水量、pH的测定分别采用烘干法、玻璃电极法;有机质测定采用重铬酸钾氧化-外加热法;全氮测定采用凯氏定氮法;全磷测定采用NaOH熔融-钼蓝比色法;全钾测定采用NaOH熔融-火焰光度法;有效磷测定采用NaHCO3浸提-钼蓝比色法;碱解氮测定采用扩散法。
1.3 土壤线虫的分离与鉴定称取土壤新鲜样品30.0 g,浅盘法室温分离48 h[13],用孔径为0.045 mm的筛子收集线虫,用5%甲醛溶液固定线虫。在解剖镜下进行计数,制片。利用土壤含水量的测定结果,将线虫个体数量转换成条·100 g-1干土。在体视显微镜下观察,将线虫鉴定到属。
各属优势度的划分:个体数量占总捕获量10%以上者,为优势属(+++);个体数量占总捕获量1%~10%者,为常见属(++);个体数量占总捕获量1%以下者,为稀有属(+)[14]。根据食性将线虫分为如下几个营养类群:食细菌性线虫(Bacterivores)、食真菌性线虫(Fungivores)、植食性线虫(Plant-parasites)和杂食/捕食性线虫(Omnivores /predators)[14]。
土壤线虫群落结构分析采用以下指标:
香农-威尔多样性指数(Shannon-Weiner index):
| $ {H^\prime } = - {n_{\rm{i}}}/N \times \ln \left( {{n_{\rm{i}}}/N} \right) $ |
Pielou均匀度指数:
| $ {J^\prime } = {H^\prime }/\ln S $ |
辛普森优势度指数(Simpson index):
| $ \lambda = \Sigma {\left( {{n_{\rm{i}}}/N} \right)^2} $ |
Margalef丰富度指数:
| $ {\rm{SR}} = (S - 1)/\ln N $ |
成熟度指数(Maturity index,MI,不包括植食性线虫)[15]:
| $ {\rm{MI}} = \Sigma c(i) \times {p_{\rm{i}}} $ |
植物寄生线虫指数(Plant parasite index,PPI)[15]:
| $ {\rm{PPI}} = \Sigma c(i) \times {p_{\rm{i}}} $ |
线虫通路比值(Nematode channel ratio,NCR):
| $ {\rm{NCR}} = {\rm{Ba}}/({\rm{Ba}} + {\rm{Fu}}) $ |
式中,ni为第i类群的个体数,N为群落所有类群的个体总数;S为类群数;c(i)为非植物寄生性(植物寄生性)土壤线虫第i类群colonizer-persister值;n为非植物寄生性(植物寄生性)土壤线虫类群数;pi为土壤线虫群落非植物寄生性(植物寄生性)土壤线虫第i类群的个体数占群落总个体数的比例;Ba、Fu分别代表食细菌性线虫、食真菌性线虫的数量。
1.4 数据处理采用IBM SPSS Statistics22.0软件进行单因素ANOVA分析和线性相关分析,以估测样地不同深度土层以及土壤理化性质对土壤线虫群落各参数的影响;用Excel绘制图表及计算。土壤理化性质与土壤线虫群落的关系采用CANOCO 4.5软件进行RDA分析及作图。
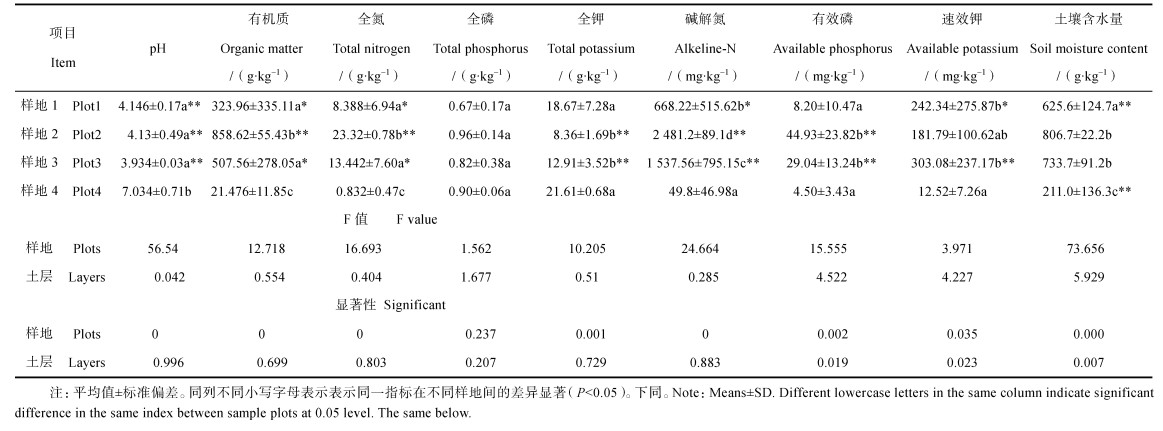
2 结果 2.1 土壤化学性质不同样地的土壤理化性质如表 2所示。位于河边滩地的样地4与其他3个样地、尤其是样地3之间的差异较明显,pH、有机质、全氮、全钾,以及速效养分均达到了显著(P < 0.05)甚至极显著水平(P < 0.01),全磷在4个样地间无明显差异。
|
|
表 2 土壤理化性质 Table 2 Soil physical and chemical properties relative to sample plot |
从4个样地共获得土壤线虫345条,隶属于5目29科34属。线虫个体密度平均278条·100 g-1干土,介于0~1 231条·100 g-1干土之间。具体情况见表 3。方差分析结果显示,土壤线虫个体密度在样地间的差异不显著(P=0.249 > 0.05,F=1.563);0~5 cm土层与20~25 cm土层之间差异较大(P=0.030 < 0.05)。总体来说,各样地的土壤线虫个体密度在土层间无明显差异(P=0.109 > 0.05,F=2.390)。土壤线虫个体数量的空间分布情况如图 1所示。
|
|
表 3 土壤线虫群落组成及c-p值 Table 3 Composition and c-p values of the soil nematode communities |

|
图 1 土壤线虫个体数量的空间分布 Fig. 1 Spatial distribution of number of soil nematode individuals |
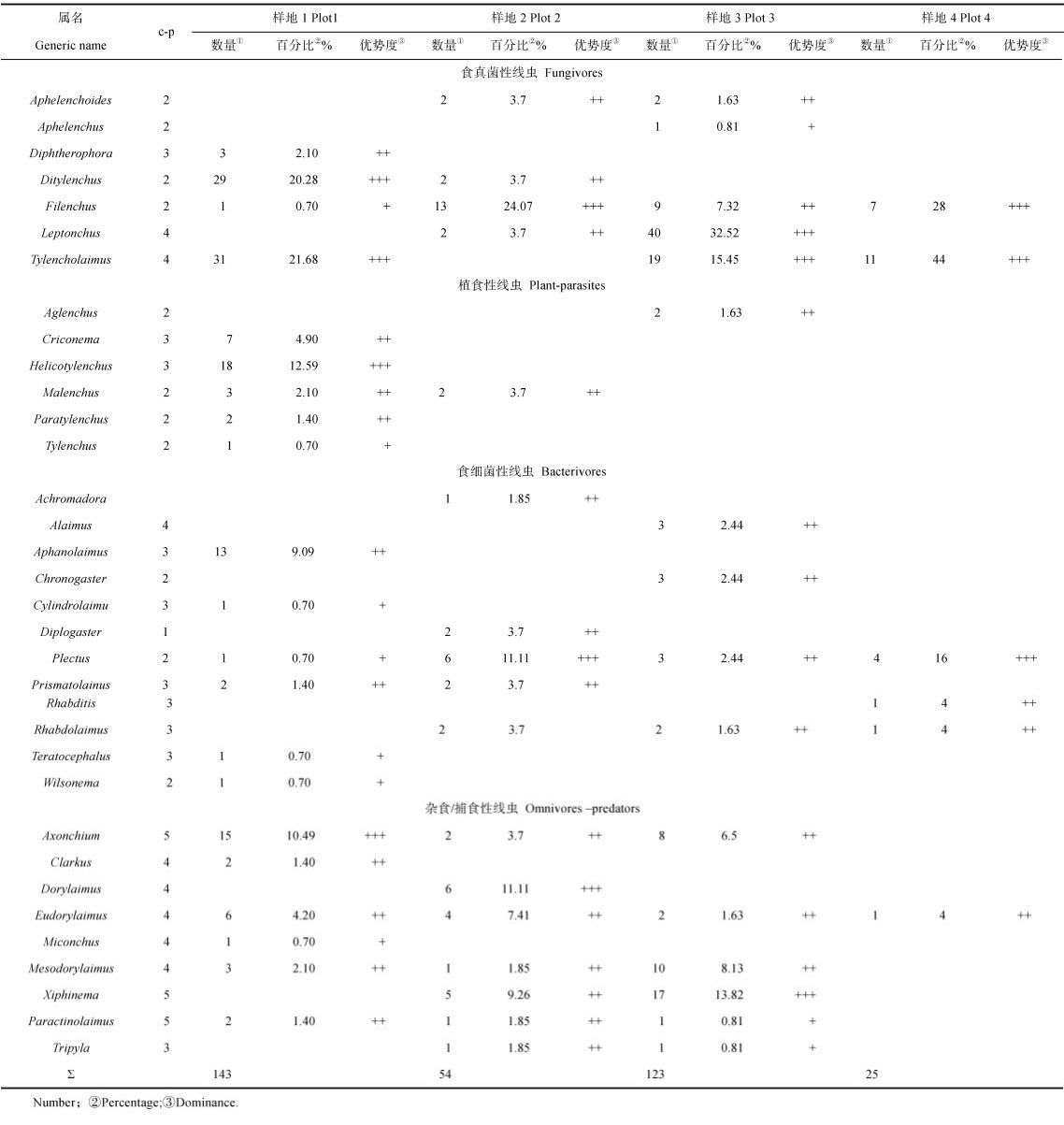
样地1获得土壤线虫21属。细齿属Leptonchus、Ditylenchus、螺旋属Helicotylenchus、Axonchium为优势属,分别占群落线虫个体数量的21.68%、20.28%、12.59%、10.49%。食真菌性线虫占44.76%,植食性线虫21.68%,杂食/捕食性线虫20.28%,食细菌性线虫13.29%。
样地2获得土壤线虫17属。Filenchus、Plectus、矛线属为优势属,分别占群落线虫个体数量的24.07%、11.11%、11.11%。食真菌性线虫和杂食/捕食性线虫分别占35.19%,食细菌性线虫25.93%,植食性线虫3.70%。
样地3获得土壤线虫16属。细齿属Leptonchus、垫咽属、Xiphinema为优势属,分别占群落线虫个体数量的32.52%、15.45%、13.82%。食真菌性线虫占57.72%,杂食/捕食性线虫占30.89%,食细菌性线虫9.76%,植食性线虫1.63%。
样地4获得土壤线虫5属。细齿属Leptonchus、Filenchus、Plectus为优势属,分别占群落线虫个体数量的44.00%、28.00%、16.00%。食真菌性线虫占72.00%,食细菌性线虫24.00%,杂食/捕食性线虫占4.00%,植食性线虫0.00%。
总体而言,垫咽属Tylencholaimus、细齿属Leptonchus的个体数量分别占到线虫总数的17.68%和12.17%,是嘎隆拉铁杉林土壤线虫群落的优势属。食真菌性线虫是主要营养类群,个体数量占到总数的49.85%,其次是杂食/捕食性线虫,占25.80%,植食性线虫所占比例最低,仅10.14%。
2.4 土壤线虫群落c-p类群组成样地1中,c-p1~c-p5类群在群落中所占的百分比分别为0%、26.57%、31.47%、30.07%、11.89%;样地2中各类群所占百分比分别为3.70%、46.30%、11.11%、24.07%、14.81%;样地3中分别为0%、16.26%、2.44%、60.16%、21.14%;样地4中分别为0%、44.00%、8.00%、48.00%、0%。
c-p1类群仅在样地2出现,数量极少;c-p2类群在样地2和样地4几乎占到1/2,数量较大。c-p3~c-p5类群在不同样地中均占有较大的比重。将四个样地作为整体,c-p3~c-p5类群占到72.17%,为优势类群。
2.5 土壤线虫群落生物多样性采用多样性指数H′、均匀度指数J′、优势度指数λ及丰富度指数SR来表征铁杉林不同样地土壤线虫群落的生物多样性(表 4)。
|
|
表 4 土壤线虫群落生态指数 Table 4 Ecological index of soil nematode community |
多样性指数H′介于0~2.31之间,平均1.34;均匀度指数J′与介于0.6~1之间,平均0.85;优势度指数λ介于0.13~1之间,平均0.39;丰富度指数SR介于0~3.37之间,平均1.95。
根据LSD和Duncan检验结果,样地4的多样性指数H′显著低于样地1、2、3(P1<0.01,P2、3<0.05),优势度指数λ有刚好相反的趋势;均匀度指数J′与样地1、2、3之间的差异均达到极显著水平(P1、2、3<0.01);丰富度指数SR,样地4与样地1、2之间有明显差异(P1<0.01,P2<0.05),与样地3则差异不明显(P3 > 0.05)。
采用成熟度指数MI、植物寄生性线虫指数PPI、线虫通道指数NCR来表征铁杉林土壤线虫群落的功能结构特征。
4个样地不同土层的MI值介于2.35~4.00之间,平均3.59。样地3的MI值最高,其次是样地1,样地2明显低于样地1、3(P1<0.05,P3<0.01)。样地4仅有3个土层分离出线虫,且数量太少,数据不计入统计。
4个样地不同土层的PPI值介于0.00~3.00之间(注:有9个样本未能分离获得植物寄生性线虫,PPI=0.00,占样本总数的50%,另外9个样本的PPI值介于2.00~3.00之间),平均值1.25。样地1的PPI值远高于其他3个样地(P2、3、4 < 0.01)。
NCR值介于0.00~0.78之间,平均0.23。除了样地2的0~10 cm层和样地1的20~25 cm层,大多数样本的NCR值均低于0.5。
2.6 土壤线虫群落与土壤性质之间关系--RDA分析及相关性分析采用冗余分析(RDA)进一步探讨土壤化学性质对土壤线虫群落的影响,分析结果见图 2及表 5。

|
图 2 土壤线虫群落与土壤性质间的RDA分析 Fig. 2 RDA of the soil nematode community with soil properties |
|
|
表 5 RDA排序结果 Table 5 Redundancy analysis collation |
RDA分析表明,土壤化学性质与土壤线虫数量之间关系密切,9个环境变量解释了77.7%的数据总差异,其中第一轴和第二轴的解释率分别为77.7%和0%,物种-环境因子排序轴的相关系数分别为0.881和0.808,物种-环境关系的累积百分比达100%,说明第1个排序轴的物种-环境相关系数很高,解释了物种和环境总方差的100%,排序图较好地反映了土壤线虫群落与土壤性质之间的关系(表 5)。
如图 2所示,粗箭头代表土壤性质各因子,小箭头代表了土壤线虫群落各指数。粗箭头越长表示某一土壤性质因子对土壤线虫群落的影响越大。箭头连线和排序轴夹角表示某一土壤因子与排序轴相关性大小,夹角越小相关性越高。
速效钾(AK)、有机质(OM)、全氮(TN)、水解氮(HN)、全磷(TP)、含水量(MC)、有效磷(AP)与土壤线虫群落的个体密度(ID)、均匀度指数J′、丰富度指数SR,以及植物寄生性线虫指数PPI和线虫通道指数NCR呈正相关;pH、全钾(TK)与上述各群落指数均呈负相关,仅与优势度指数λ和成熟度指数MI呈正相关关系。
根据箭头长短,速效钾(AK)、有机质(OM)、全氮(TN)、全钾(TK)对土壤线虫群落的影响较大,是主要影响因子;根据夹角则显示出,速效钾(AK)对PPI的相关性最强,其次是J′和个体密度(ID);全钾(TK)与MI有较好的相关性。
并非每个土壤性质因子对土壤线虫群落均有显著影响。如表 6所示,应用线性相关性检验分析每个土壤性质因子对线虫群落影响的显著性,土壤速效钾对线虫群落个体密度、多样性变化以及功能指数均有显著影响(P个体密度、PPI < 0.01;PH'、λ、SR、MI < 0.05);土壤含水量对线虫群落个体密度、多样性,以及MI有显著影响(P个体密度、、λ、MI < 0.05;PH'、SR < 0.01);土壤pH对线虫群落多样性及MI有显著影响(PH'、SR < 0.01;PMI、λ < 0.05);土壤有机质、全氮、水解氮、有效磷对线虫群落个体密度有显著影响(P < 0.05);有机质、全氮对SR有显著影响(P < 0.05);有效磷对H'有显著影响(P < 0.05);全磷、全钾、水解氮对MI有极显著影响(P < 0.01)。
|
|
表 6 土壤线虫群落与土壤性质的相关系数 Table 6 Correlation coefficient between soil nematode community and soil properties |
本实验所设置的四个样地代表了嘎隆拉山环境条件基本一致但林分类型有所差异的四种铁杉林类型。实验共鉴定到土壤线虫345条,隶属于5目29科34属,个体密度介于0~1231条∙100 g-1干土之间,平均278条∙100 g-1干土。
2008年10月,凌斌等[16]在高黎贡山海拔2 878 m处云南铁杉针阔混交林调查到的土壤线虫个体密度低于120条∙100 g-1干土,种类大约34属(文中未提供准确数据,读图获得大致的数值)。本研究与凌斌等的调查采样工作均在秋季进行,高黎贡山云南铁杉针阔混交林土壤线虫个体密度远低于嘎隆拉山铁杉林,而线虫属数则大致相同。同属藏东南亚高山暗针叶林的色季拉山急尖长苞冷杉林,在海拔约3 800 m处分离获得土壤线虫41属,个体密度平均406条∙100 g-1干土。嘎隆拉山铁杉林土壤线虫无论数量还是种类均低于同地区的色季拉山急尖长苞冷杉林。生态指数H′、SR也均低于色季拉山急尖长苞冷杉林。根据冗余分析和线性相关性分析结果,线虫个体密度与土壤含水量之间极显著正相关,因此,水分分配格局或许是导致上述不同生境土壤线虫个体密度产生差异的重要原因之一。
此外,嘎隆拉铁杉林土壤线虫群落的优势属是垫咽属Tylencholaimus和细齿属Leptonchus,食真菌性线虫是最主要的营养类群。这个结果与色季拉山急尖长苞冷杉林土壤线虫群落以垫咽属、螺旋属及绕线属为优势属,植食性线虫是主要营养类群的结果不同[7],表现出嘎隆拉铁杉林土壤线虫群落种类构成对环境的适应性,以及功能上的变化。
四个样地中,样地4的土壤理化性质与其他样地差异较大,线虫群落的个体数量、属数均低于其他样地,其种类构成也表现出了一定的不同。这个样地位于河滩地,接近铁杉林在本区域分布的上限,为天然更新的幼树林,草本盖度远低于其他三个样地,pH明显升高,有机质含量以及速效氮磷钾则明显偏低,土壤化学性质出现明显不同。生境的贫营养及pH值的增高导致样地4土壤线虫群落生物多样性显著低于其他样地。样地4在线虫多样性降低的同时,统计结果显示其个体密度与其他样地间并没有明显差异,这或许与群落结构中c-p值为1~2的r-生存策略者所占比重较大(44%)有关。生境条件的劣化会导致r-生存策略者通过大量繁殖来替代在该环境条件数量下降或缺失的线虫种类,以维持群落相对稳定。这与Walker[17]所提出的“冗余种”假说一致,表现出该样地土壤线虫在生存策略上的特殊性。
c-p3~c-p5类群为k-生活策略者,它们生活周期长,增殖能力低,对干扰敏感,在生境中稳定而持续地生活的时间较长,当土壤被扰动时,有可能导致其数量的下降或缺失。而作为土壤线虫典型机会主义者的cp1类群和机会主义者cp2类群,生命周期短,增殖能力强,耐受干扰、富营养化和厌氧条件,在有利条件下数量会明显增加[18],而当环境变劣时也会通过大量繁殖来应对环境的变化。总体而言,嘎隆拉山铁杉林土壤线虫群落以c-p3~c-p5类群为主要线虫种类,表明嘎隆拉山铁杉林生态系统受到的干扰较小,处于较为稳定的状态。该区域所受干扰主要来自于自然因子,例如水热分配变化,这点在样地4得到了体现。成熟度指数MI平均值为3.59,与同地区色季拉山急尖长苞冷杉林3.65(平均值)的调查结果[7]相近,证明嘎隆拉山铁杉林生态系统所受干扰较小,处于较为稳定的状态[15]。
PPI值则与扰动的频率呈正相关[19]。嘎隆拉山铁杉林的PPI值远小于色季拉山急尖长苞冷杉林2.66的调查结果,表明嘎隆拉山铁杉林被扰动的频率远低于色季拉山急尖长苞冷杉林。四个样地中,样地1的PPI值远高于其他3个样地,由此推测该样地受到扰动的频率高于其他样地。
NCR值低于0.5的研究结果显示,嘎隆拉山铁杉林的枯落物主要时通过真菌降解途径为主,这与色季拉山急尖长苞冷杉林土壤线虫的调查结果一致[8]。
冗余分析(RDA)结果说明了土壤性质与土壤线虫群落之间的关系情况。速效钾、有机质、全氮、全钾对土壤线虫群落的影响较大,是主要影响因子,其中,速效钾是土壤化学性质中最为重要的影响因子。从测试结果可以看出,嘎隆拉山铁杉林土壤有机质、速效氮、磷、钾含量均属于高肥力水平,在此条件下,速效钾所表现出的重要性值得关注。相关研究中[20-22]针对碳、氮的成果较多,针对钾的相对较少,本研究关于钾的重要性与张荣之等[23]在贡嘎山不同垂直气候带土壤线虫群落分布格局的研究结果一致。线性相关分析结果说明速效钾显著影响线虫群落的数量和多样性,而有机质、全氮、水解氮、有效磷则是通过影响线虫数量从而影响着群落结构。冗余分析(RDA)结果表明,土壤含水量与土壤线虫群落的个体密度、均匀度指数J′、丰富度指数SR,以及植物寄生性线虫指数PPI和线虫通道指数NCR均呈正相关关系,线性相关分析结果也说明了土壤含水量对线虫群落个体密度、多样性,以及MI均有显著性影响,由此说明环境水分条件对土壤线虫群落的种类构成及数量有着不可忽视的影响。此结果表明,不同气候带间线虫群落密度及多样性均无显著差异,但食真菌、捕食-杂食和植物寄生线虫的密度差异显著,土壤有效氮、全钾及温度是主要的影响因子。
4 结论嘎隆拉山铁杉林土壤线虫隶属于5目29科34属,个体密度介于0~1 231条∙100 g-1干土之间,平均条∙100 g-1干土;垫咽属Tylencholaimus和细齿属Leptonchus为优势属;食真菌性线虫是最主要的营养类群;枯落物主要通过真菌途径进行降解。冗余分析(RDA)及线性相关分析结果表明,土壤速效钾、有机质、全氮、全钾是土壤理化性质中对土壤线虫群落影响较大的几个因子,其中,速效钾最为重要;线虫个体密度与土壤含水量之间极显著正相关,水分分配格局或许是导致嘎隆拉山铁杉林不同生境土壤线虫个体密度产生差异的重要原因之一。嘎隆拉山铁杉林生态系统受到干扰较小,处于较稳定状态。
| [1] |
Liu L H, Qiu X Z. Studies on the geographic distributions and situations of vertical zone of the Chinese Tsuga forest (In Chinese)[J]. Acta Botanica Yunnanica, 1980, 2(1): 9-21. [刘伦辉, 邱学忠. 我国铁杉林的地理分布及垂直带位置的研究[J]. 云南植物研究, 1980, 2(1): 9-21.]
(  0) 0) |
| [2] |
Chen L Z, Chen Q L, Liu W H. Forest diversity and its geographical distribution in China (In Chinese). Beijing: Science Press, 1997. [陈灵芝, 陈清郎, 刘文华. 中国森林多样性及其地理分布[M]. 北京: 科学出版社, 1997.]
( 0) |
| [3] |
Liu J S, Zhang B, Zhou X W, et al. Establishment of biomass density model of spruce forest in Tibet (In Chinese)[J]. Forest Resources Management, 2018(1): 38-43. [刘金山, 张蓓, 周学武, 等. 西藏自治区云杉林生物量密度模型研建[J]. 林业资源管理, 2018(1): 38-43.]
( 0) |
| [4] |
Shen Z Q, Lu J, Hua M, et al. Point pattern analysis of five main plant populations of dark coniferous forest in the Sejila Mountains, Tibet, China (In Chinese)[J]. Journal of Northwest Forestry University, 2016, 31(1): 7-14. [沈志强, 卢杰, 华敏, 等. 西藏色季拉山暗针叶林5个主要植物种群点格局分析[J]. 西北林学院学报, 2016, 31(1): 7-14.]
( 0) |
| [5] |
Li J R, Fang J P, Lu J, et al. Analysis on point patterns of two main plant populations in subalpine dark coniferous forest of southeast Tibet (In Chinese)[J]. Forest Resources Management, 2014(1): 104-108, 119. [李江荣, 方江平, 卢杰, 等. 藏东南亚高山暗针林2个主要植物种群点格局分析[J]. 林业资源管理, 2014(1): 104-108, 119.]
( 0) |
| [6] |
Xie C Q, Tian M X, Zhao Z R, et al. Spatial point pattern analysis of Abies georgei var. Smithii in forest of Sygera Mountains in southeast Tibet, China (In Chinese)[J]. Chinese Journal of Applied Ecology, 2015, 26(6): 1617-1624. [解传奇, 田民霞, 赵忠瑞, 等. 西藏色季拉山急尖长苞冷杉种群点格局分析[J]. 应用生态学报, 2015, 26(6): 1617-1624.]
( 0) |
| [7] |
Chen Y, Yang W Q, Wu F Z, et al. Diversity of soil nematode communities in the subalpine and alpine forests of western Sichuan, China (In Chinese)[J]. Chinese Journal of Applied Ecology, 2017, 28(10): 3360-3368. [谌亚, 杨万勤, 吴福忠, 等. 川西亚高山/高山森林土壤线虫多样性[J]. 应用生态学报, 2017, 28(10): 3360-3368.]
( 0) |
| [8] |
Xue H Y, Luo D Q, Yu B Z. Community characteristics of soil nematode in Abies georgei var. smithii forest in Sejila Mountain of Tibet, Southwest China (In Chinese)[J]. Chinese Journal of Applied Ecology, 2012, 23(12): 3402-3408. [薛会英, 罗大庆, 于宝政. 西藏色季拉山急尖长苞冷杉林土壤线虫群落特征[J]. 应用生态学报, 2012, 23(12): 3402-3408.]
( 0) |
| [9] |
Liu X, Xu Z H, Yu N N, et al. Ant species diversity of mount Galongla and Medog valley in southeastern Tibet (In Chinese)[J]. Forest Research, 2017, 30(1): 34-40. [刘霞, 徐正会, 于娜娜, 等. 藏东南嘎隆拉和墨脱河谷蚂蚁群落研究[J]. 林业科学研究, 2017, 30(1): 34-40.]
( 0) |
| [10] |
Bongers T, Ferris H. Nematode community structure as a bioindicator in environmental monitoring[J]. Trends in Ecology & Evolution, 1999, 14(6): 224-228.
( 0) |
| [11] |
Xu J J, Zhao B, Zhang L Y, et al. Assessment of genetic diversity of Rhododendron nivale in Tibetan, China using AFLP markers (In Chinese)[J]. Bulletin of Botanical Research, 2017, 37(1): 88-95. [徐静静, 赵冰, 张良英, 等. 西藏地区雪层杜鹃遗传多样性的AFLP分析[J]. 植物研究, 2017, 37(1): 88-95.]
( 0) |
| [12] |
Lu R K. Analytical methods for soil and agro-chemistry (In Chinese). Beijing: China Agricultural Science and Technology Press, 1999. [鲁如坤. 土壤农业化学分析方法[M]. 北京: 中国农业科技出版社, 1999.]
( 0) |
| [13] |
Mao X F, Li H X, Chen X Y, et al. Extraction efficiency of soil nematodes by different methods (In Chinese)[J]. Chinese Journal of Ecology, 2004, 23(3): 149-151. [毛小芳, 李辉信, 陈小云, 等. 土壤线虫三种分离方法效率比较[J]. 生态学杂志, 2004, 23(3): 149-151.]
( 0) |
| [14] |
Wu D H, Yin W Y, Bu Z Y. Changes among soil nematode community characteristics in relation to different vegetation restoration practices in the moderate degraded grasslands of Songnen (In Chinese)[J]. Acta Ecologica Sinica, 2008, 28(1): 1-12. [吴东辉, 尹文英, 卜照义. 松嫩草原中度退化草地不同植被恢复方式下土壤线虫的群落特征[J]. 生态学报, 2008, 28(1): 1-12.]
( 0) |
| [15] |
Bongers T. The maturity index:An ecological measure of environmental disturbance based on nematode species composition[J]. Oecologia, 1990, 83(1): 14-19. DOI:10.1007/BF00324627
( 0) |
| [16] |
Ling B, Xiao Q M, Ge F, et al. Composition and diversity of soil nematode community in Gaoligong mountains in Yunnan Province (In Chinese)[J]. Journal of Hunan Agricultural University:Natural Sciences, 2008, 34(3): 341-346. [凌斌, 肖启明, 戈峰, 等. 云南省高黎贡山土壤线虫群落结构及多样性[J]. 湖南农业大学学报:自然科学版, 2008, 34(3): 341-346.]
( 0) |
| [17] |
Walker B H. Biodiversity and ecological redundancy[J]. Conservation Biology, 1992, 6(1): 18-23. DOI:10.1046/j.1523-1739.1992.610018.x
( 0) |
| [18] |
Bongers T, Bongers M. Functional diversity of nematodes[J]. Applied Soil Ecology, 1998, 10(3): 239-251. DOI:10.1016/S0929-1393(98)00123-1
( 0) |
| [19] |
Ruess L. Nematode soil faunal analysis of decomposition pathways in different ecosystems[J]. Nematology, 2003, 5(2): 179-181. DOI:10.1163/156854103767139662
( 0) |
| [20] |
Wang M W, Liu Y D, Chen X Y, et al. Response of soil nematode community to cultivation in upland red soil relative to cultivation history and its signifi cance as indicator (In Chinese)[J]. Acta Pedologica Sinica, 2016, 53(2): 510-522. [王明伟, 刘雨迪, 陈小云, 等. 旱地红壤线虫群落对不同耕作年限的响应及指示意义[J]. 土壤学报, 2016, 53(2): 510-522.]
( 0) |
| [21] |
Wang X X, Gao Q Z, Hasbagan Ganjurjav, et al. Soil nematode community response to warming in alpine meadow of northern Tibet (In Chinese)[J]. Pratacultural Science, 2018, 35(6): 1528-1538. [王学霞, 高清竹, 干珠扎布, 等. 藏北高寒草甸土壤线虫群落结构对增温的响应[J]. 草业科学, 2018, 35(6): 1528-1538.]
( 0) |
| [22] |
Zhang M, Liang W J, Zhang X K. Soil nematode abundance and diversity in different forest types at Changbai Mountain, China[J]. Zoological Studies, 2012, 51(5): 619-626.
( 0) |
| [23] |
Zhang R Z, Liu X L, Zhong H M, et al. Distribution pattern of soil nematode communities alongthe vertical climate zones on the eastern slope of Gongga Mountain (In Chinese)[J]. Chinese Journal of Applied and Environmental Biology, 2016, 22(6): 959-971. [张荣芝, 刘兴良, 钟红梅, 等. 土壤线虫群落在贡嘎山东坡不同垂直气候带间的分布格局[J]. 应用与环境生物学报, 2016, 22(6): 959-971.]
( 0) |